The Descent of English: New Evolutionary Models of Language Change
Daniel Newman © 2008
1. Introduction
2. Language evolves?
3. A new evolutionary paradigm
4. Applying evolutionary theories to changes in English
5. Conclusion
6. Glossary
7. Works cited
1. Introduction: Language Changes
Anyone with even the most casual interest in language will know it is not static. Parents complain about teenage lexicons, and inevitably teenagers grow up to deplore the way kids speak these days. Our English is different from Dickens’s English and even more so from Shakespeare’s. Chaucer, in what seems like a different language, recognizes and simultaneously demonstrates that language changes:
Ye know ek that in forme of speche is chaunge
Withinne a thousand yeer, and wordes tho
That hadden pris, now wonder nyce and straunge
Us thinketh hem, and yet thei spake hem so
– Chaucer, Troilus and Criseyde II.22 – 25 (ca. 1380)
This article reviews recent developments in linguistics, which seek evolutionary explanations for language change. It is divided into three parts. Section 2 gives a brief history of evolutionary thinking in linguistics. Section 3 outlines some recent historical linguistic work toward a theory of language change based on Darwinian principles. Finally, Section 4 shows some of the potential applications of language-evolutionary theories to changes observed in the history of English.
1.1. Why Study Language Change?
Mutability complicates the study of any subject; moreover it can be greatly disturbing to essentialist or ideal conceptions of what that subject is. As Roger Lass writes, humans have long recognized the universality of change, and have often contrasted it with “an ‘ideal’ stasis” (278). Darwin’s theory of descent by modification was revolutionary because it challenged the Scriptural account of Creation, in which all species are immutable (Dennett 65).
Thus linguistic purists resist loanword adoption, neologisms, and the vicissitudes of pronunciation. Such conservatism is often associated with Joseph Addison, Jonathan Swift and other 18th-century notables (Beal 17–18), but it survives today, often among newspaper and blog pundits. Certainly it is easier to understand an unchangeable thing than one continually becoming new things. It is easier to speak of English if one believes, in theory at least, that there is such thing as the English language.
In reality languages are ever-changing. They change over historical time; between generations; across geographical distances; and even within a speaker’s lifetime. Therefore language change has been a central concern in linguistics since the emergence of the field in the nineteenth century. Its causes, manifestations, and dynamics remain primary problems for contemporary linguists (Ritt 2004:55–56).
2. Language Evolves?
Given the temporal changeability of language, it is tempting to say it evolves. There are senses in which this is not controversial. We might say quite reasonably that languages evolve from older and into newer ones (OED v6, 8, 9). It is difficult to speak of evolution, however, without evoking the specific case of biological evolution—which in the post-Darwinian era usually means evolution by natural selection.
2.1. Analogies between Biological Evolution and Language Change
Many affinities between languages and organisms make metaphorical transfer appealing. Indeed, like living systems, languages
· change over time;
· transmit information from generation to generation;
· give rise to new forms and go extinct;
· form internally-intelligible groups (e.g., English ≈ Homo sapiens) made up of unique individuals (i.e., idiolects);
· are formed by variously-defined subsets divided by geography and other factors (e.g., dialects ≈ subspecies);
· involve “grammars” (i.e., language competence ≈ genome) and “texts” (i.e., acoustic and graphical representation ≈ phenotype).
(Adapted from Ritt 2004:89–90.)
2.2. A Brief History of Language Evolution
These affinities were not lost on early linguists. Indeed, the links between biology and linguistics date back to the 19th century. In fact, an exchange of ideas and metaphors between scholars of language and natural history helped both disciplines grow into their modern forms (Croft 2000:9; Fitch 665).
Because linguistic systems are transformed organically over time, Darwinian models seem well-suited for describing language change. Relatedness among languages and among organisms has long shared metaphors of genealogy, using family trees to represent relationships among biological taxa and linguistic groups. Biology and linguistics independently developed cladistics to reconstruct statistically these family trees (Lass 113–114; Dunn et al. 2072–2073; see McMahon & McMahon’s Language Classification by Numbers).
As early as 1543, Theodor Bibliander described Hebrew and Arabic as related languages based on their similar scripts, lexis, and morphologies (Lass 108). 19th-century philologists borrowed explicitly from Darwin’s theories (Fitch 665), some going as far as considering languages as living organisms (Ritt 2004:55). However, by the early 20th century such practices had fallen out of favour (McMahon 1994:314). Early language evolutionists were too hasty in their enthusiasm to apply Darwinism to language; thus their models were merely metaphorical, bearing little relation to actual changes undergone by languages (Ritt 2004:56).
2.3. Difficulties with Evolutionary Theories of Language Change
William Croft claims that evolutionary theories of language change often fail “because of improper analogies” (2004:73). Although useful for generating theory and guiding research questions (Sereno 468), analogical thinking can be misleading. Furthermore, if linguists hope to formalize a truly evolutionary theory—one which explains how and under what conditions language actually changes—metaphorical transfer is not sufficient (Foster 19; Ritt 2004:89–91).
Evolutionary theories of language change have repeatedly met impasses, where biology ceases to elucidate linguistic processes. Salikoko Mufwene names the transmission of information as a most problematic discrepancy between biological and linguistic evolutionary models. Whereas offspring inherit all their genes at once from their one or two parents (in asexual or sexual reproduction, respectively), humans learn language from innumerable sources over their lifetime (Mufwene 47). Language is “genetic” in the sense that information is transmitted in discrete packets like words. However, language change is also Lamarckian, meaning that language-traits acquired during an individual’s lifetime can then be transmitted to others (McMahon 1994:336; Mufwene 46). Organisms do change in their lifetimes (e.g., bigger muscles from exercise), but these modifications are not transmitted to their offspring.
Thus the problem of transmission complicates what seem like convincing analogies between biology and language. Consequently, 20th-century linguists tended to dismiss evolutionary models (McMahon 1994:314). Nevertheless, recent developments in biology, neuropsychology, and Complexity Theory have reignited interest in language evolution. In Explaining Language Change, April McMahon (44–45; 47–48) argues for new theories to explain the problems of sound change actuation and transmission; her cautious conclusion is that these theories should be evolutionary (336).
3. A New Evolutionary Paradigm for Language Change
Ideally, an evolutionary theory of language change would uncover the simple set of principles that underlies the complex dynamics of language change. As such, it could unify the field of linguistics (Ritt 85; McMahon 1994:138–139; Worden 353), just as Darwinism forms the basis for all modern biology (Dennett 147).
Like McMahon, Roger Lass advocates an evolutionary theory of language change. He suggests a medium-neutral evolutionary model, which sees language “neither in any individual…nor in the collectivity, but rather as an area in an abstract, vastly complex, multi-dimensional phase-space” (376–377; cp. Labov’s phonological-space graphs: 4.4.1.1 ). Historical linguistics would benefit from considering speakers not as “‘language builders’…, but end-users of historically evolved systems of a certain kind, which share many properties with other such systems” (xviii). Language and its users should be viewed separately. Lass is cautious about using evolutionary approaches too literally, but his arguments anticipate those of later, explicitly Darwinian, theorists. His contention that language can and ought to be seen apart from human minds is primarily methodological, but it also suggests his conceptual affinities with Selfish Gene Theory. McMahon raises the same point when she describes “speakers of a language as ‘blind watchmakers’, fitting pieces into a pattern they cannot see” (1994:337).
3.1. Rationales for Language-Evolution Models
By virtue of being past, the past is difficult to reconstruct reliably. The imperfect and more or less random survival of data constitutes one of the great challenges for historical linguistics (Labov 11). Missing links in the fossil record embarrassed Darwin’s analysis of macroevolutionary patterns; thus gaps in and unreliability of much of the historical linguistic evidence make past language change difficult to reconstruct.
However, if past linguistic changes obey general evolutionary rules, then the same rules likely drive contemporary changes, which are easier to observe. Insofar as general principles can be isolated from contemporary data, present trends may provide insight into historical ones for which evidence is scant. Thus a Darwinian theory of language change potentially addresses the “Historical Paradox,” which states that “to the extent that the past was different from the present, there is no way of knowing how different it was” (Labov 21). If a Darwinian processes underlie language change, then it is reasonable to infer past language changes from ongoing ones.
3.2. A New Approach: Generalised Darwinism
Darwinism explains how life evolved and continues to evolve. Recently, a growing number of historical linguists have reenergised the field of evolutionary linguistics by suggesting that language evolves in accordance with generalised Darwinian principles. These concepts of language assume that some linguistic unit operates as a replicator, like a gene, and that replicators act in certain general ways known as complex adaptive behaviour (Mufwene 47).
3.2.1. Language as a Complex Adaptive System (CAS)
Various natural and cultural phenomena qualify as complex adaptive systems (CAS). These systems exhibit complicated and even chaotic dynamics, but nevertheless change and self-organize according to a simple set of general principles (Ritt 2004:91–93). Biological life is the most familiar CAS, but artificial life, human and animal societies, stock markets, and national economies may also be understood as complex systems that exhibit evolutionary dynamics (Ritt 2004:58). Recent language evolutionists posit that language also changes according to these simple, underlying processes (e.g., Mufwene 47). This is not to say that languages evolve by Darwinian evolution. Life is only one particular CAS among many. Instead, recent language evolutionists speak of “generalised Darwinism” as a model for language change. A set of underlying general principles govern the evolutionary behaviour of all CAS, each of which has its particularities. Finding what these principles are is the major challenge for language evolutionists.
3.3. The Requirements for Evolvability
It is important, then, to isolate the fundamental processes required for Darwinian evolution. Biology provides the basic requirements for evolvability thus:
a. There must be variation among individuals within interacting populations. Variation provides the raw materials for evolution by natural selection. For a population to evolve, variation must be already present within it.
b. The variation must be heritable, inheritance occurring through self-replication of the unit of information (i.e., a gene).
c. There must be differential propagation of types dictated, on average, by the relative advantages conferred by inherited variants. Popularly and misleadingly known as “survival of the fittest.”
Evolution occurs when the relative proportion of types in a population changes. Consider a population of Eastern Grey Squirrel (Sciurus carolinensis), a species with a fur-colour gene that makes individuals either grey or black. If an original population of 40 % black and 60% grey squirrels is later found to be composed of 45% black and 55% grey, evolution has occurred. In extreme cases, one type reaches 100% of the population.
In biological systems, the unit of inheritance is the gene—a small bit of DNA coding for a specific protein. What I have called a type above is in fact a gene-type, or genotype.
3.3.1. Example of Biological Evolution: Squirrel-Fur Colour
The graphic below shows in more detail the evolution of fur colour in a hypothetical population of squirrels.
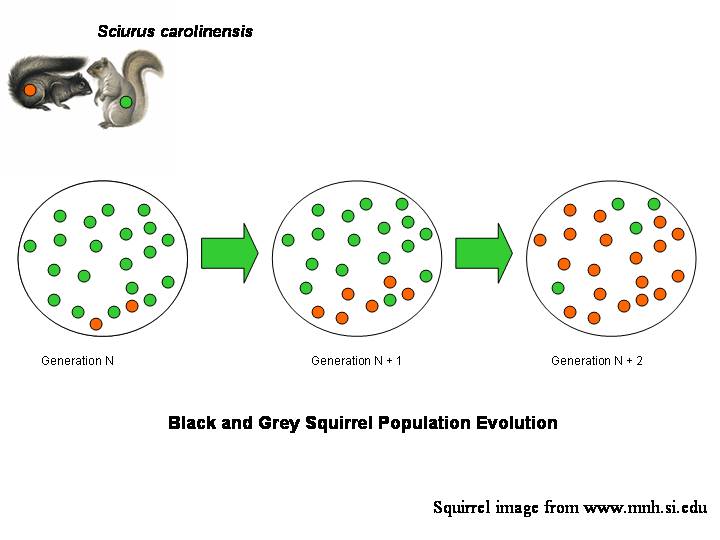
Figure 1. Cartoon of three generations of an evolving population of Eastern Grey Squirrel. There are two genotypes, grey and black fur, represented with green and orange circles respectively. Thus the genotype can be considered separately from the individual animals (i.e., vessels) that carry them.
The grey type, which is dominant in Generation N, is gradually displaced and then almost entirely replaced by the black type by Generation N + 2. Changes in relative proportion may be due to selective advantage (e.g., house cats prefer eating grey types, or black types are less prone to heart attacks); to chance (e.g., a tree falls on a large gathering of grey individuals); or, more likely, to a combination of advantage and chance.
The next cartoon shows the same dynamics occurring in the primary population, but includes a second, insular population, in which the grey-fur type survives alone.
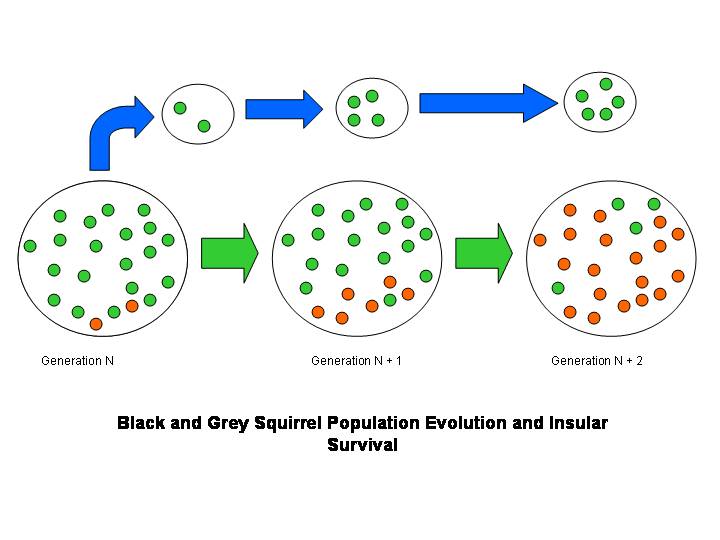
Figure 2. The same population as in Figure 1, but with a migration event by individuals carrying the grey-fur gene in Generation N. None of the migrants have the black-fur gene; therefore the breakaway population does not undergo the same fur-colour evolution—even if exposed to the same selection pressures (which favour black-furred individuals). There is simply no variation for selection to act on.
3.4. Applying Darwinism to Language
We may now apply these basic Darwinian principles to language:
a. Variation. Everyday experience amply demonstrates that language is variable. Differences occur at the level of lexicon, pronunciation, syntax, etc. They are cultural, geographical, generational, and idiosyncratic.
b. Heritability. Things get more complicated here. Like genes, language is clearly passed on from parents to their children, but other people and media also contribute to language acquisition. Salikoko Mufwene specifies that whereas a genotype “may…be said to be transmitted wholesale, a language is not” (47). The elements of language are not inherited faithfully, but rather adopted into personalized idiolects (47). By contrast, genetic information is passed from parent to offspring with remarkably few errors. However, this is a difference of degree rather than of kind. The fact that humans inherit language throughout their lives, and from multiple sources, does not challenge the generalised-Darwinian model. Still, the linguistic version of inheritance is considerably more complex than the genetic one. The real problem with the heritability requirement is figuring out what a language gene actually is. We shall return to this problem shortly.
c. Differential propagation. As we have seen in the squirrel example, initial variation may be whittled down to one type. The same is true of language. For example, multiple early spellings of the loanword barbecue (i.e., barbecu, borbecu, barbicue, barbacue, and babracot [OED]) were eventually lost, leaving only the contemporary variant. Conversely, variation may spur diversification; historical variation in the spelling of en/insure has over time begotten two different words, ensure and insure (OED). Selective advantage may be conferred to one variant over another by means of grammar guides, dictionaries, social shibboleths, nationalistic revival, etc. Chance plays a role too: one may imagine a typo in a bestseller that leads to a new, accepted word.
3.4.1. Example of Language Evolution: /iː/ in Bite
Figure 3 adapts the squirrel example for language, replacing fur-colour with pronunciation of the áiñ in the word bite. (N.B. Pronunciation is only one example of the possible loci of language evolution; alternatively, the figure below might show changing patterns of lexis [e.g., sweat versus perspire], of verb-forms [e.g., dived versus dove], etc).

Figure 3. Cartoon of three generations of an evolving population of the phoneme /i:/ in the word bite. Here three types are shown, whereas the squirrel example had two; however, the principle is the same.
The cartoon below reproduces the conditions of the second squirrel cartoon. As in Figure 2, a variant displaced by other types persists in an isolated breakaway population. Here the formerly-dominant [iː] pronunciation of bite survives in an insular population, although it is almost extinct in the original one.
The lack of Cockney /h/-dropping in North American English exemplifies this pattern. In England, it /h/-dropping from words like heart and heat (thus /ɑːt/ and /iːt/) originated in the 18th century, but only after the majority of British emigration to North America. Therefore, /h/-dropping is almost absent in North America, while it has spread widely throughout the United Kingdom (Nevalainen 2000:38; Wells 255).

Figure 4. Same original phoneme population as in Figure 3, but with a migration event in Generation N, and the establishment of a non-variable breakaway population of [iː]. Compare with Figure 2.
3.5. Analogies, Homologies, Conceptual Difficulties
The examples above show some intriguing correspondences between biological and language change. However, many difficulties frustrate the development of an effective evolutionary theory. There is, for example, the non-trivial task of determining the relative contributions of language-internal and -external factors (Farrar & Jones 1). In additions, various theories are difficult to reconcile and compare because their conceptual parameters differ significantly; for example, some see words as linguistic species (e.g., Worden 353), and others as the equivalents of genes (e.g., Croft 2000: 27–28).
For these reasons, recent theoreticians have focused on clarifying the conceptual and methodological frameworks required in a well-organized, intelligible research program. The challenge for language evolutionists is to discover in language functional, empirically-observable homologues (as opposed to analogues) of biological units and processes. Table 1 shows some of the many intriguing links exist between biological species and linguistic languages; between biological populations and linguistic communities; and between an organism’s genome and a speaker’s grammar.
Table 1. Some correspondences between biology and linguistic units and processes. (Adapted from Croft 2000:38, with some revisions or simplifications. Examples are mine.)
Generalised theory |
Biological evolution |
Language evolution |
Example of language evolution |
replicator |
gene |
language gene |
/i/ |
replicator population |
gene pool |
language-gene pool |
English language in hypothetical Pop’n X |
replicator sequence |
DNA molecule |
utterance |
“bite” |
faithful replication |
reproduction (sexual or not) |
communication (utterance production) |
“bite” ® “bite” |
replication error |
recombination, mutation |
innovation miscommunication |
“bite” ® “bite me” “bite” ® “buy it” |
replicator variants |
alleles (e.g., fur-colour gene comes in two alleles, grey and black) |
variants |
In Pop’n X, /i/ comes in two variants, [i:] and [aI] |
vessel (interactor) |
organism (body) |
speaker (including her grammar) |
Betty |
environment |
ecological context |
socio-communicative context |
Betty’s familial, social, cultural, educational, and professional surroundings |
selection regime |
survival and reproduction |
longevity and communication of language conventions among speakers |
Generational preference for “bite me” seen as uncool by new generation, and loses competition to more popular expression. |
3.5.1. Hunting for the Replicator
The linguistic equivalent of a gene is the source of the greatest difficulty and controversy in language-evolution theories. Without it, all the higher-level affinities are merely analogical. This is no trivial problem, because truly evolutionary processes require that changes occur by replication (Croft 2006:73). That is, in order to fit the requirements of the generalised Darwinism model, a system must exhibit change through the iterative but imperfect production of copies (Croft 2006:73; Dawkins 1992:25; Dennett 48–60; Ritt 2004:122–124).
3.5.1.1. The (Selfish) Language Gene.
In theory, this is how a language gene might spread through a speech community:
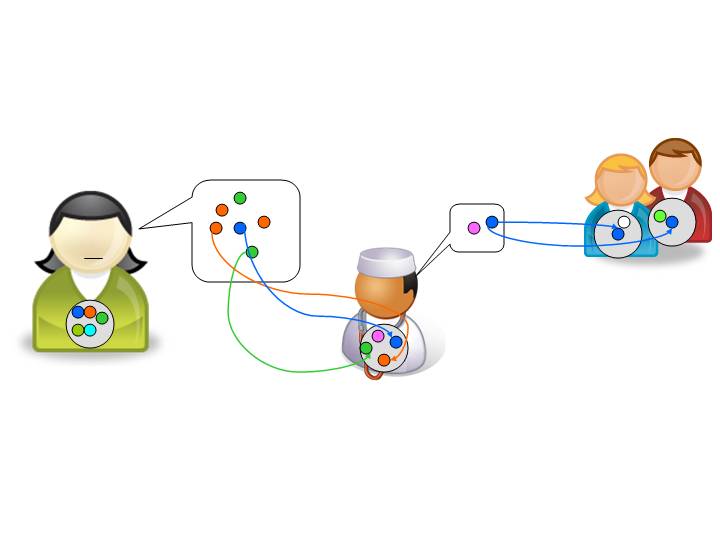
Figure 5. A cartoon representation of selfish language-gene transmission. Each speaker has an individual grammar (grey circles), equivalent to a genome, which contains all her language genes (coloured circles within the grammar). When these language genes are expressed as utterances, they can be incorporated into the grammar of hearers; hearers, in turn, may pass on the language genes (e.g., the trajectory of the blue language gene from left to right). (Adapted from Croft 2000:27–28.)
3.5.1.2. Qualities of a Linguistic Replicator
Language genes as we currently understand them are not readable. Genes are. As physical information-coding objects (i.e., segments of DNA molecules), they can be read by chemical analysis and other means. What a language gene is and how it might be observed remain unclear. As McMahon cautions over-enthusiastic language evolutionists, “We do not yet know which units of selection might operate in historical linguistics; are they words, rules, speakers, or languages themselves?” (337).
Darwinism can help. In evolutionary theory, language evolutionists find on the fundamental qualities of linguistic replicators, which include:
· materiality (i.e., they must be detectable empirically);
· enough stability to permit transmission among speakers; and
· enough copy-faithfulness to make transmission intelligible.
(Adapted from Croft 2000:27–28.)
Much theoretical work by language evolutionists has therefore gone into finding what constitute a linguistic replicator. And yet McMahon’s statement still applies after almost 15 years of theoretical exploration and speculation.
3.5.2. A Few Language-Replicator Hypotheses
Roger Lass (1997) proposes one of the earliest replicator-based language-evolution models. William Croft (2000) and Nikolaus Ritt (2004) follow with monographs predominantly concerned with language replicators (linguemes according to Croft, memes to Ritt). The models differ in important ways, but share a fundamental belief in generalised Darwinism.
3.5.2.1. A Quasi-Species Model of Language Evolution
Language replicators—whatever they may be—copy themselves with far less accuracy than genes do. Therefore Lass bases his model on the quasispecies (375–376), “a well-defined distribution of mutants that is generated by a mutation-selection process” (Nowak 118). The quasispecies concept was designed to explain the evolutionary dynamics of viruses like HIV, which replicates itself extremely quickly and with extremely high rates of replication error (Nowak 120).
A hypervariable virus population is probably the closest biological equivalent to a language population. In the mutation-selection process, a single variant, occupying a single coordinate in phonological space will begin to replicate itself. (For phonological space, imagine a Cartesian graph of two, three, or more dimensions, where a coordinate is the intersection of the x, y, z,….nth axes). Because the variant replicates itself imperfectly, it forms a “cloud” of mutant “offspring” around itself. Small mutations are more likely than big ones, and big mutations may fall into inhospitable areas of phase-space. Therefore the density of mutants is higher near the original replicator. However, a very different mutant may arise in a particularly suitable area in phonological space; thus its own “offspring” will begin to form a dense cloud around it. These relatively stable areas of high-density are quasispecies, and are the targets of selection.
The quasispecies model is particularly applicable to language evolution because it deals with hypervariable replication, as one would expect from linguistic genes. Thus the word bite is never spoken exactly the same way, even by a single speaker, and its associative and symbolic relationships continually adjust themselves among and within individuals. As long as the word is intelligible to hearers, its variants (e.g., /baIt/, /baIət/, /beIt/) are viable. But extreme variations on pronunciations of bite like /bɒt/ and /bIt/ may become confused with boat and bit, respectively, and thus be eliminated by selection.
Lass finds the model useful, too, because “it allows us to concentrate on our subject matter [i.e., language], and not worry about the hardware (or the software) too much” (377). In other words, the model focuses on populations of linguistic units rather than populations of speakers using linguistic units.
3.5.2.2. William Croft’s Utterance-Based Model
According to Croft, the language gene is a linguistic structure embodied in an utterance. It take be a phoneme, morpheme, word, syntactic construction, etc—any linguistic structure actually used by a speaker (2000:28).
In the utterance-selection model, language genes are replicated through a speaker’s utterances. The survival of these replicators require that the speaker survives long enough to produce it in speech, therefore transmitting it to hearers, who will then be able to utter them. Altered replication, required for the production of variants upon which selection can act, is effected by the inevitably intra- and interindividual differences in utterance (Croft 2000:38).
3.5.2.3. Nikolaus Ritt’s Neural Network Hypothesis
Ritt objects to Croft’s model, which treats utterances as separate from their interpretation in the minds of speakers and listeners (158). Instead proposes a much more complicated type of neural replicator, which considers both utterances (“texts” in his words) and the linguistic competences involved in producing and understand them. The replicator is, in his words, “an assembly of nodes in a network of neurally implemented constituents,” whose size, structural and, ultimately, longevity depend on its being used in associative cognitive activity (169). For example, the language gene that codes for the morpheme anti- will develop a complex and well-used neural network because it is a commonly-used affix with a wide range of applications; in contrast, the morpheme pygo- (relating to the buttocks; OED) is too rare in most circles to result in such a well-developed network of neurons.
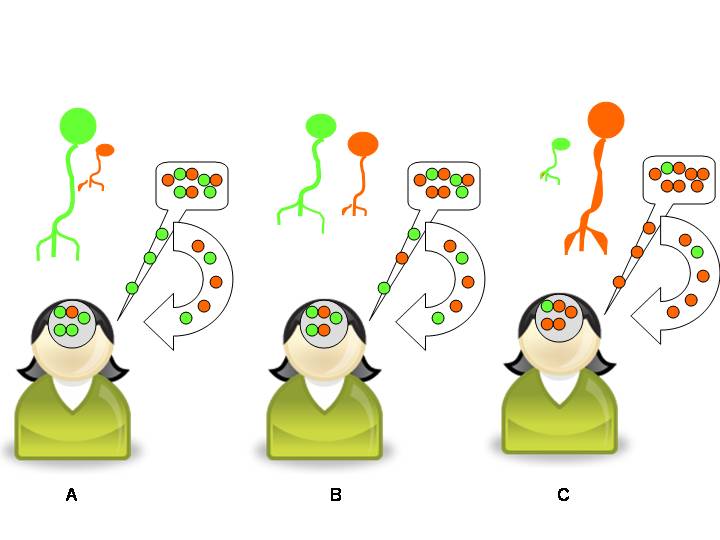
Figure 6. Schematic simplification of Ritt’s neural-network model. The speaker’s “grammar” (or language competence) is the grey circle drawn on her head. The green and orange circles in it represent two variant types of language gene (for example, two pronunciations of bite). More frequently-used variants form more developed neural networks (represented by the squid-like nerve cells above the speaker). In A, the speaker’s neural network reflects greater use of the green variant, although she is increasingly using the orange on in speech (see the speech bubble in A). Her language use feeds back into her mind (see the curved arrow from the speech bubble to her head). This gradually alters the relative proportion of green and orange variants in her mind, thus changing the neural networks for the pronunciation of bite. In B, the orange network has grown with use, while the green one has become to give way. In C, the orange variant has almost totally displaced the green variant. Compare with Figures 3 and 4.
3.5.2.4. Comparing Croft’s and Ritt’s Replicator Models
Croft’s theory has the advantage of simplicity, but Ritt’s objections are embarrassing. Similarly, Mufwene disagrees with Croft’s equation of the language gene with the utterance, arguing that “Speakers do not learn to reproduce other speakers’ utterances. Instead they learn (some of) the units and principles that enable those speakers to produce utterances” (47n).
Ritt’s own theory is a much less elegant working model, and will have to wait long for neurological research to detect anything like his neural replicators. Ritt’s replicator has the additional advantage of being a material entity and therefore observable by empirical methods. It has the major disadvantage of being entirely speculative, reaching far beyond current neurology. This is not a problem per se, but it does make application of the model somewhat problematic. More importantly, it is not clear how Ritt's neural networks fit the requirements of replicators. What is replicated in this model is not the neural network itself; the network is no more than a template or mould for storing and processing language information. That is, it does not carry information itself; nor can it be transmitted from brain to brain. Clearly, neural networks would play a role in any complete “genetic” theory of language, but in my opinion that role is not the equivalent of DNA.
Nevertheless, such hypotheses provide more and more compelling and integrated theory. Until replicators are found, however, the theory remains metaphorical. This is not to say that language-evolutionary thinking cannot be useful in tackling actual problems in historical linguistics. Applying evolutionary models to changes in the history of English is thus subject of following discussion.
4.1. Applying Evolutionary Models to Patterns of Change in the History of English
English, with its well-studied history, dialectal diversity, and geographical distribution, is an excellent model for demonstrating and questioning the claims of linguistic evolutionists. Its lineage is relatively well known, and much work has been done on its development and variability. As a colonial language, it has adopted new forms worldwide, which can be viewed as settler populations with more or less independent post-contact evolutions. As a global language, it continues to evolve in parallel but interacting lineages.
Before turning to my primary example of language evolution, which focuses on chain shifts in vowel pronunciation, I briefly examine two other aspects of English evolution. Two very recent articles in Nature use evolutionary theory and statistics to study lexical and grammatical innovation and conservation over time. As such, they represent significant empirical applications of Darwinism in historical linguistics, and may help refine and guide models like those advocated by Croft, Ritt, and others.
4.1.1. Lexical Evolution and Word-Use Frequency
Word use is correlated with lexical conservatism. Mark Pagel et al. compare language corpora of English, Russian, Spanish, and Greek, as well as 200 “fundamental vocabulary meanings” in almost a hundred Indo-European languages, and find that the more a given word is used, the less vulnerable it is to being replaced. Word-use frequency accounts for 50% of the variation in the rates of lexical replacement. Thus oft-used English words like two, tongue, and die have remained remarkably similar to their cognates in other Indo-European languages, whereas less frequent words like stab, dirty, and guts have diverged (717).
The study claims to confirm the suitability of evolutionary models in studies of language change. Because frequent words tend to outcompete or suppress the emergence of new synonymous lexemes, they may be conserved for tens of thousands of years. Despite the apparent ethereality of linguistic units, “the purifying force of spoken word frequency… can have a replication accuracy as high as that of some genes” (719); therefore, common words and their durability provide concrete support for the “culturally transmitted replicator” (719) required by generalised-Darwinian theories.
4.2.2. Grammatical Evolution and Irregular-Verb Usage
Use is crucial determinant of lexical survival. As Erez Lieberman and colleagues demonstrate, it also has grammatical implications. Tracking the loss of strong-verb forms in English, they find that of 177 Old-English verbs, only 145 and 98 remain irregular in Middle and Modern English respectively (713). As in Pagel’s study, frequency of use is inversely related to the rate of change. Extremely common irregular verbs like to be, to have and to go are least likely to adopt the regular –ed past-tense form, while rarer ones like to wring, to wed and to swell have not yet but are most likely to become regular (714–715). Lieberman et al. present the tendency of irregular verbs to become regularized as a competitive dynamic, where more powerful grammatical rules outcompete more complicated or unwieldy ones (715). As the number of regularized verbs increases, the likelihood that other irregular verbs will also be regularized grows; this is because more speakers will apply the –ed rule by analogy as the number of already-standardized verbs grows. However, selection will preserve irregular verbs in their so-called primitive states, as long as speakers use them frequently enough not to analogically subject them to the –ed rule.
4.3. Past: The Great Vowel Shift (GVS)
A contentious event in linguistics provides an excellent arena for testing new approaches (McMahon 2006:9). The Great Vowel Shift (GVS), a major change in the way long vowels were pronounced in English (Nevalainen 2006:120), is the subject of persistent controversy. Linguists disagree on whether the chain shift resulted from a “push” caused by the raising of low and low-mid vowels, or from a “pull” caused by the diphthongization of the high vowels /iː/ and /uː/ (McMahon 2006:6–7; Nevalainen 2006:122). Some linguists question whether a “great” vowel shift occurred at all, viewing the various sound-changes as more or less independent rather than concerted events (McMahon 2006:7).
Language-evolutionary thinking may help to resolve such debates. At the very least, it may guide linguists toward hypotheses about how changes associated with the GVS might have occurred.
4.3.1. The Push- versus Pull-Chain Debate
A tentative evolutionary analysis of the GVS supports the Push-Chain theory. The merging of vowels like /eː/ and /iː/ can cause competition between them (McMahon calls it “pressure” [2006:8]). Competition is especially acute where sound mergers result in homophony, as with meet and meat. When the áañ in mate is raised to [e], thus merging with áeañ in meat, the sounds compete for the same phonological space (see Labov 266). This in turn pushes the áeañ in meat to up to [eː]. Ambiguity between closely-merged vowels can lead the members of a speech community to favour variants of /ea/ that are noticeably different from the raised /a/. Thus competition between merging vowels can selectively favour further sound changes. By contrast, the diphthongization of the high vowels /iː/ and /uː/ to [eI] and [ou] would not cause such competition, as diphthongization pushes pronunciation into unoccupied phonological space.
From an evolutionary perspective, a push shift seems more parsimonious than a pull shift. The push-shift scenario offers a mechanism for sequential vowel-sound changes (i.e., competition for phonological space), which a pull shift lacks. This is not to suggest the problem is as simple as I present it; moreover, there is no reason to think the initial sound-change in a chain shift cannot be toward unoccupied phonological niche. The analysis above is more than anything a highly simplified but plausible illustration of how language evolutionists might address historical-linguistic controversies like the GVS.
4.3.2. Pronunciation Variation: Raw Materials for the Great Vowel Shift
The Great Vowel Shift may be approached as an evolutionary change in English pronunciation (Samuels 144–153). Thus, sound changes associated with the GVS are spurred by a selective regime favouring some vowel variants over others. There must have been a time when some speakers pronounced bite as /biːt/ while others said /baIt/. Then, for whatever reason, the latter variant more effectively spread through English-speaking communities, becoming the standard pronunciation
Where did these vowel variants come from? In fact, the vowel variation necessary for sound competition and phonetic evolution is ubiquitous in real speech communities. Although it is impossible to get a clear picture of pronunciation in England at the time of the GVS, it was probably no less diverse than it is today (Ritt 2004:42–44).
The sound [iː] in green provides an excellent example. Although /griːn/ is the Standard-English pronunciation, the 1978 Linguistic Atlas of England by Orton, Sanderson & Widdowson (cited by Ritt 2004:43–44) reveals extensive geographical variation in contemporary England. While [iː] dominates in most of the country, there are small areas in the south where [eː] is predominant; large ones in the north where [ei] or [əi] is most widespread; and enclaves were [iə] prevails locally (Ritt 2004:42–44). It is unlikely that any speech community uses only the dominant form; it seems reasonable to assume that several variants coexist with it in each territory.
· Variation. There is geographical variation in the pronunciation of green, which includes /greːn/, /grein/, griən/ in addition to /griːn/.
· Heritability. Regional variation despite normative forces (e.g., media, increased travel/migration) suggests that local socio-communicative transmission can be remarkably faithful.
· Differential propagation. Where /greːn/ persists, one could say that the /eː/ lingueme outcompetes rival variants—even /iː/.
If we assume, like Ritt, that geographical and individual variation vowel pronunciation is not a new phenomenon—i.e., it occurred at the time of the GVS—we find the conditions for a selfish-gene evolutionary shift. A vowel variant gains selective advantage and spreads, thus forcing out the previous dominant sound (Ritt 2004:43 – 48).
/griːn/ is standard today because the language gene [iː] replicated more successfully than other language genes. Why this occurred is more difficult to determine, but we could imagine various causes: the rise to political or social prominence by members of a group for whom /griːn/ was the native pronunciation; or the wild popularity of a poem that rhymes green with sin (/siːn/).
Ritt’s green example highlights how evolutionary processes might work on existing variation. However, the exact historical context that gave [iː] selective advantage over, say, [eː] cannot be determined. His example is also highly simplified. It views a single vowel shift without considering concurrent changes in other sounds. Such reductionism is necessary for the purposes of illustration. However, more empirical data are necessary for better representation of observed historical changes.
4.3.3. Using Historical Data
Some historical data do survive. This permits at least some empirical analysis of past language change. Rhyming poetry and dictionaries have proved important data-sources for historical pronunciation (Nevalainen 2006:118–119). In addition, some insight into pronunciation-change is may be gained from historical spellings (Ritt 2004:45). For example, spellings of the word green has varied in interesting ways:
pre-1000s–1100s: gróeni, gréne
1200s–1700s: grene
1400s–1600s: grenn, grenne, greyn, greyne
1400s–1700s: greene, gren
1500s–present: green
1600s: greane, grein, greine, gryne
1700s: grien (OED “green, a. and n.”).
The appearance of áiñ and áyñ in 17th-century spellings of green may reflect the spread of [iː], which would eventually dominate its pronunciation (Ritt 2004:45).
However, such data provide only limited vision of past patterns. Furthermore, their reliability is difficult to ascertain. For this reason, language evolutionists must seek more complete sources of data.
4.4. Using the Present to Understand Historical Changes
Studying language change in the past has its benefits. For one, large-scale patterns are often detectable only from a historical remove. But then again, these patterns may be spurious artefacts, borne from the incomplete survival of documents and from the pattern-seeking by linguists (Lass 38; Nevalainen 2006:120). Furthermore, inevitable gaps in the record make it difficult to develop an empirically-workable theory of language evolution; indeed, the less empirical evidence there is, the more researchers must resort to generalization, extrapolation, and theorization (Labov 11). For this reason, language-evolutionary studies have a better chance of finding underlying principles by observing contemporary language changes.
4.4.1. The Northern Cities Shift
The Northern Cities Shift (NCS) is a major vowel shift currently underway in some urban centres in the Northeast United States (Labov 23). Three decades of research into the phenomenon provide extensive data (Ito 3), from which linguists like William Labov seek to find the general rules of chain shifting (116–121).
Labov notes that though his empirical approach differs in method and philosophy from the evolutionary theories, each seeks “to find the most general principles that govern language structure and language change” without contradicting the other (115). He has conducted extensive surveys of speech communities throughout the Northern Cities area (116, 177), which includes Rochester, Syracuse, Buffalo, and Chicago (Ito 4). The NCS is unique among known vowel shifts because it affects short rather than the usual long vowels (Ito 3). However, a Darwinian-inspired approach will seek the deeper phonological (≈ genetic) and social (≈ ecological) processes, which may then be applied to other instances of language change.
4.4.1.1. Microevolutionary Processes in the Northern Cities Shift
Vowel-shifts are incremental changes. There are innumerable grades between the Standard-English [æ] in bag (/bæg/) and the extreme cases of Northern-Cities’ [ε] or [I] (thus /bεg/ or /bIg/) (Ito 4). Therefore, individual-, dialectal-, and larger-level variations in the pronunciation of single vowels provide evidence that can help to refine, guide, and challenge theoretical models and reconstructions of historical language changes.
From recorded conversations, Labov produces graphical representations of how variously a single vowel-sound is pronounced by and among individuals. Thus minor variations caused by neighbouring consonants, stress, and emphasis can be represented as a data-cloud in two- or multi-dimensional phonological space (165n, 174). Figure 7 represents a hypothetical phonological-space graph of the sound /iː/ in green.

Figure 7. Hypothetical graph showing an individual’s variable pronunciation of /ee/ in the word or prefix green or green-. “grèen” represents green in a secondary-stress position; “green” in a primary-stress position; “green!” in an emphatic-stress position. The two axes represent two different phonological variables. (Based on Labov 174.)
By comparing such graphs for individuals from the same, adjacent, and distant areas, researchers can track geographical, temporal, and idiosyncratic variation in pronunciation. With such information, it may be possible to extract some of the microevolutionary rules that ultimately contribute toward larger patterns of pronunciation changes.
4.4.1.2. Possible Analogies with the Quasispecies Model?
A multi-dimensional phonological graph like the one above can be made of individual- as well as speech-community-level variation. Roger Lass’s linguistic quasispecies model may be applicable to study how clouds of neighbour variants might be selected over others. Thus a “cloud” of neighbouring /ee/ pronunciations may be shown undergoing selection at the speech-community level and migrating in phonological space. (The analogy is suggestive, but whether it has practical applications is beyond the scope of this author.)
5. Conclusion: Data and Theory, Challenges and Promise
The earliest attempts to describe language as a complex adaptive system are not yet two decades old. Whether generalised Darwinism proves as revolutionary for linguistics as Darwinism did in biology remains to be seen. Evolutionary theories of language change are still immature, still unsure of their parameters.
Empirical research such as Labov’s is crucial to the development of a workable theory of language evolution. Likewise, verbal and mathematical models like those by Croft, Ritt, and Mufwene are critical for finding general principles from disparate empirical studies, and for guiding data collection. While many of these theories reach beyond the scope of current empirical research, they will in time be vindicated by evidence or replaced by new theories that better explain it.
This is not to downplay the achievement of the research programme or the usefulness of its findings and theories. Indeed, attempts to push historical linguistics into a paradigm shift will surely reveal much about the structure of language and the ways in which it changes. It is also important to remember that nearly a hundred years passed between the publication of The Origin of Species and the discovery that DNA is the molecule that carries genetic information. During this interval great advances were made based only on the hypothetical notion of a unit of heredity (Worden 354–355). Perhaps the future of language evolution holds such breakthroughs.
6. Glossary
Genome: “The total genetic makeup of an organism” (Rudin 162). It is the complete assemblage of genes an individual inherits from its parents, whether or not these genes are expressed. It is more or less equivalent to a speaker’s grammar, her total knowledge of her language, whether used or not (Croft 2000:28).
Genotype: The specific gene-variants carried by an individual, whether expressed or not (Rudin 162).
Language evolution (in contrast with evolutionary linguistics): I use language evolution to differentiate between the study of languages as evolving systems and evolutionary linguistics, the study of language as part of the evolved structure of the human mind (i.e., work by Steven Pinker, Noam Chomsky, and others).
Language gene: Throughout the article, I use the general term language gene for the still-undiscovered unit of linguistic replication. Some authors prefer more specific terms: meme (Ritt 2004:120; Worden 354; after Dawkins 1976:206) or lingueme (Croft 2000:28). Although language gene most obviously refers to the biological gene, it is important to note that the units of linguistic replication exist at various levels of linguistic order, ranging from morphemes to syntactic constructions (Mufwene 47).
Macroevolution: Major evolutionary change resulting in new taxonomic groups (e.g., species, orders) (Rudin 217). The emergence of the English, German, Dutch, Frisian, and Flemish languages from West Germanic (McMahon & McMahon 3) is a macroevolutionary event.
Micorevolution: Genetic change that occurs within populations…as a result of progressive adaptation” (Rudin 228) and random genetic drift (Rudin 160). A linguistic equivalent is the pronunciation changes associated with the Great Vowel Shift. Microevolution over evolutionary time leads to macroevolution.
Phenotype: The expression of genes in an individual (Rudin 284).
Quasispecies: Martin Novak summarizes a quasispecies thus:
A quasispecies is a well-defined distribution of mutants that is generated by a mutation-selection process. Selection does not act on a single mutant but on the quasispecies as a whole…. A quasispecies may be centered around a master [genetic] sequence with high efficiency of reproduction. The consensus sequence of the quasispecies need not be identical with the master sequence…. The quasispecies has changed the classical view of evolution. (118–121).
The OED defines it as:
a group of similar but non-identical DNA or RNA molecules, with a distribution centred on a hypothetically ideal sequence, resulting from a balance between mutation and selection pressure. In later use also: a population of viruses with genomic sequences of this nature. (OED n2.)
Selfish Gene Theory: Richard Dawkins’ selfish gene theory is the current paradigm in evolutionary biology. It views the body as a vessel constructed by genes to better their chances of spreading. Successful genes are those that make fit bodies for themselves; fit bodies survive and reproduce better than less fit ones, and thereby pass more copies of their genes into the next generation. In other words, the body is ephemeral and dies, but the genetic and language-genetic codes it has carried may live on, provided they are passed on successfully. Obviously, genes are not actually selfish, as they lack any conscious will or agenda. Instead, differential survival over time makes it look as though successful genes were acting selfishly; in fact, they do not survive because they are selfish, but appear selfish because their ability to propagate themselves allow them to survive. See Dawkins 1976 or Dennett 1995 for detailed discussions of selfish genetics.
7. Works Cited
“barbecue, n.” The Oxford English Dictionary. 2nd ed. 1989. Oxford UP. 31 Dec. 2007. http://dictionary.oed.com.myaccess.library.utoronto.ca/cgi/entry/50017490.
Beal, Joan C. English in Modern Times. New York: Oxford University Press, 2004.
Chaucer, Geoffrey. “Troilus and Criseyde.” The Riverside Chaucer.
Croft, William. Explaining Language Change: an Evolutionary Approach. Toronto: Longman, 2000.
Croft, William. “Evolutionary Models and Functional-Typological Theories of Language Change.” The Handbook of the History of English. Eds. Ans van Kemenade and Bettelou Los. Maldan MA: Blackwell Publishing, 2006. 68–91.
Dawkins, Richard. The Selfish Gene. Toronto: Paladin, 1976.
Dawkins, Richard. “Universal Biology.” Nature 360 (1992): 25–26.
Dennett, Daniel. Darwin’s Dangerous Idea: Evolution and the Meanings of Life. New York: Touchstone Books, 1995.
Dunn, Michael, Angela Terrill, Ger Reesink, Robert A. Foley, and Stephen C. Levinson. “Structural Phylogenetics and the Reconstruction of Ancient Language History.” Science 309 (2005): 2072–2075.
“Eastern Grey Squirrel, Sciurus carolinensis.” Smithsonian Natural Museum of Natural History. n.d. 4 Jan. 2008. http://www.mnh.si.edu/mna/image_info.cfm?species_id=298.
“ensure, v.” The Oxford English Dictionary. 2nd ed. 1989. Oxford UP. 3 Jan. 2008. http://dictionary.oed.com.myaccess.library.utoronto.ca/cgi/entry/50076028.
.
“evolve, v6, 8.” The Oxford English Dictionary. 2nd ed. 1989. Oxford UP. 24 Dec. 2007. http://dictionary.oed.com.myaccess.library.utoronto.ca/cgi/entry/50079227.
“evolve, v9.” The Oxford English Dictionary. Additions Series 1993. Oxford UP. 24 Dec. 2007. http://dictionary.oed.com.myaccess.library.utoronto.ca/cgi/entry/50079227.
Farrar, Kimberley & Mari C. Jones. “Introduction.” Eds. Mari C. Jones & Edith Esch. Language Change: the Interplay of Internal, External and Extra-Linguistic Factors. New York: Mouton de Gruyter, 2002. 1 – 18.
Foster, Michael. “Language Change and Darwinian Principles.” Change and Language. Ed. Hywel Coleman and Lynne Cameron. Philadelphia: Multilingual Matters, 1996.
“green, a. and n.” The Oxford English Dictionary. 2nd ed. 1989. Oxford UP. 3 Jan. 2008. http://dictionary.oed.com.myaccess.library.utoronto.ca/cgi/entry/50098386.
“insure, v.” The Oxford English Dictionary. 2nd ed. 1989. Oxford UP. 3 Jan. 2008. http://dictionary.oed.com.myaccess.library.utoronto.ca/cgi/entry/50118480.
Ito, Rika. “Diffusion of Urban Sound Change in Rural Michigan: a Case of the Northern Cities Shift.” Diss. Michigan State University, 1999.
Labov, William. Principles of Linguistic Change: Volume 1: Internal Factors. Cambridge: Blackwell, 1994.
Lieberman, Erez, Jean-Batiste Michel, Joe Jackson, Tina Tang, Martin A Nowak. “Quantifying the Evolutionary Dynamics of Language.” Nature 449 (2007): 713 – 716.
McMahon, April. “Change for the Better? Optimality Theory versus History.” The Handbook of the History of English. Eds. Ans van Kemenade and Bettelou Los. Maldan MA: Blackwell Publishing, 2006. 3–23.
McMahon, April. Understanding Language Change. Cambridge: Cambridge UP, 1994.
McMahon, April, and Robert McMahon. Language Classification by Numbers. Toronto: Oxford UP, 2005.
Mufwene, Salikoko. “Competition and Selection in Language Evolution.” Selection 3.1 (2002): 45–56.
Nevalainen, Terttu. “Gender Differences in the Evolution of Standard English: Evidence from the Corpus of Early English Correspondence.” Journal of English Linguistics 28 (2000): 38 – 59.
Nevalainen, Terttu. An Introduction to Early Modern English. Toronto: Oxford UP, 2006.
Novak, Martin A. “What Is a Quasispecies?” Trends in Ecology and Evolution 7.4 (1992): 118–121.
Pagel, Mark, Quentin D. Atkinson, & Andrew Meade. “Frequency of Word-Use Predicts Evolution throughout Indo-European History.” Nature 449 (2007): 717 – 721.
“pygo-, comb. form.” OED Online. Dec. 2007. Oxford University Press. 3 Jan. 2008. http://dictionary.oed.com.myaccess.library.utoronto.ca/cgi/entry/50193453.
“quasispecies, n. OED Online. Dec. 2007. Oxford University Press. 4 Jan. 2008. http://dictionary.oed.com.myaccess.library.utoronto.ca/cgi/entry/50293282.
Ritt, Nikolaus. “Language Change as Evolution: Looking for Linguistic ‘Genes’.” Vienna English Working Papers 4.1 (1995): 43–56. University of Vienna English Department. 31 Dec.2007. http://www.univie.ac.at/Anglistik/vinitst.htm.
Ritt, Nikolaus. Selfish Sounds and Linguistic Evolution: a Darwinian Approach to Language Change. Cambridge: Cambridge UP, 2004.
Rudin, Norah. Dictionary of Modern Biology. Hauppauge, NY: Barron’s, 1997.
Samuels, M.L. Linguistic Evolution with Special Reference to English. New York: Cambridge UP, 1976.
Sereno, M.I. “Four Analogies between Biological and Cultural/Linguistic Evolution.” Journal of Theoretical Biology 151 (1991): 467 – 507.
Wells, John Christopher. Accents of English 1: an Introduction. Cambridge: Cambridge UP, 1986.
Worden, Robert P. “Words, Memes, and Language Evolution.” Evolutionary Emergence of Language: Social Function and the Origins of Linguistic Form. Eds. Chris Knight, Michael Stuart-Kennedy and James R. Hurford. New York: Cambridge UP, 2000.